Dendritiske celler af Langerhans. Hudens immunsystem. Mastceller og eosinofiler
CL betragtes som et udviklingsstadium af "professionelle" APC - myeloide dendritiske celler. De indtager en fundamentalt anden plads i hudens immunsystem end keratinocytter. Først kommer de ind i huden udefra, da de kommer fra myeloide celler i knoglemarven. For det andet, på trods af at CL'er modtager et aktiveringssignal i huden under immunprocesser, skal de for at udføre en antigenpræsenterende funktion forlade den og migrere til lymfeknuden.
CL er procesceller (deres nye navn ifølge den internationale klassificering er hvide procesepidermocytter), som hovedsageligt er lokaliseret i det tværstribede keratiniserende epitel, men som også findes i dermis, udgør 2-3 % af det samlede antal epidermale celler ; i alt indeholder en voksens epidermis omkring 10 9 CL.
Det morfologiske træk ved CL, ud over procesformen, er Birbeck-granulerne indeholdt i dem - tre-lags cytoplasmatiske formationer. CL klæber praktisk talt ikke til glasset, fagocyterer svagt, bestemmes i snit og suspensioner histokemisk (farvning for ATPase), samt ved at identificere karakteristiske membranmarkører. Det drejer sig primært om langerin (CD208), som er et mannosebindende C-lectin og funktionelt associeret med Birbecks granulat, samt CD68, E-cadherin, CD1a Ag og MHC klasse II molekyler (hos mennesker, hovedsageligt HLA-DR). CD4 Ag er til stede på en del af CL, men CD8 Ag, som markerer dendritiske T-celler, er fraværende. CL bærer også adhæsionsmolekyler CLA, LFA-3 (CD58), ICAM-1 (CD54), ICAM-3 (CD-50), b4-integriner, udtrykker Rts for en række cytokiner - TNF a, GM-CSF, M- KSF osv.
CL'er vises i epidermis hos mus på dag 16-17 af embryonal udvikling. Deres umiddelbare forstadier er celler af fænotypen CD14+, CD11b-, CD11c+, der cirkulerer i blodet. CL-prækursorer migrerer ind i huden på grund af tilstedeværelsen af RC CCR2 på deres overflade, som genkender b-kemokin MCP-1, som produceres af keratinocytter aktiveret under inflammation. I processen med migration fra huden til den regionale lymfeknude, under påvirkning af aktiverende signaler, omdannes LC til interdigiterende celler i lymfeknuderne.
In vitro dendritiske celler dyrkes fra en fraktion af hæmatopoietiske stamceller (fænotype CD34) eller fra monocytter (fænotype CD14) i nærvær af GM-CSF og IL-4. For varianten med dannelse af LC fra CD34-celler var nøglen introduktionen af TGF b 1 i dyrkningsmediet (sammen med GM-CSF, TNFa, stamcellefaktor og Flt3-ligand). I dets tilstedeværelse dannes LC'er indeholdende Birbeck-granulat og E-cadherin og uden det monocytter. LC dannes ud fra cirkulerende monocytter. in vitro i nærvær af IL-4, IL-10, TNFa og monoklonale AT-anti-CD40-agonister; TGF b forstærker også denne proces. Samtidig mister progenitorceller CD14 Ag og udtrykker E-cadherin og klasse II MHC-molekyler.
For de fleste af membranmarkørerne i LA er funktionen indstillet. CLA Ag-ekspression er således vigtig for CL-lokalisering i huden, E-cadherin og b-catenin sikrer etablering af LA-kontakter med keratinocytter, langerin udfører funktionen af mannosebindende RC. LA-prækursorer udtrykker ikke E-cadherin; det forsvinder ved deres aktivering, når LA forlader epidermis.
LA's evne til at udføre de indledende stadier af Ag-præsentation (binding, bearbejdning af Ag og ekspression af dets fragmenter i sammensætningen af disse molekyler) er forbundet med MHC klasse II- og CD1-molekyler. Et kendetegn ved CL er en udtalt evne til endocytose og Ag-behandling og manglen på evnen til effektivt at præsentere antigene peptider for T-hjælpere. Modne dendritiske celler i lymfeknuderne (interdigiterende celler) mister tværtimod evnen til at binde og behandle Ag, men har en udtalt evne til at præsentere det for T-hjælpere. Sidstnævnte egenskab afhænger som nævnt ovenfor ikke kun af tilstedeværelsen af MHC klasse II-molekyler, der bærer det antigene peptid på overfladen af dendritiske celler, men også af ekspressionen af costimulerende CD80- og CD86-molekyler. CL'er udtrykker CD86 svagt, men mangler CD80, mens interdigiterende celler indeholder mange costimulerende molekyler på deres overflade.
Opholdstiden for CL i huden er under normale forhold 18 måneder (i andre organer og væv - 2 måneder). I fravær af hudskader eller nogen manifestationer af biologisk aggression, afslutter CL'er deres livscyklus her. Under aktiveringsbetingelser af hudimmunsystemet, bakterieprodukter, stoffer frigivet fra beskadigede celler og lokalt dannede cytokiner (produkter af keratinocytter, T-celler og CLS selv) påvirker selv skæbnen for CLS. På grund af det faktum, at kun en vejafgiftslignende RC-TLR2 er til stede på CLS, er de i stand til at reagere på bakteriepeptidoglycaner, men ikke til lipopolysaccharider. Bakterielle peptidoglycaner, cytokiner produceret af keratinocytter, T-celler og CL selv (IL-1, TNF A, GM-CSF osv.), Såvel som midler såsom dinitrochlorobenzen og nikkelsalt, aktiverer CL. Dette udtrykkes i en ændring i membranfænotypen - udseendet på CL -membranen på CD83 -molekylet, der tjener som en markør for dendritiske celler, kemokin RC CCR7, der tilvejebringer rettet migration af CL til lymfeknuder og vedhæftning Molekyler VLA-4 og CD44, der fremmer denne migration, såvel som i en stigning i ekspressionen af MHC-molekyler. Klasser I og II, det costimulatoriske CD86-molekyle og til sidst svækkelsen af ekspressionen af E-cadherin og AG CLA , som bevarer CL i epidermis.
Aktiveret CL producerer et antal cytokiner-IL-1 A GM-CSF, interferoner samt IL-16 og B-chemokiner, der er nødvendige for at tiltrække T-celler til huden. Derudover udskiller de IL -12, hvilket er vigtigt for udviklingen af Th1 - producerende celler af IFN G og en række andre cytokiner, der er ansvarlige for udviklingen af cellulære immunprocesser i huden. Til gengæld forstærkes produktionen af IL-12 i CL af bindingen af membranmolekylet CD40 såvel som af virkningen af IFN g. GM-CSF derimod undertrykker produktionen af IL-12.
Startstimulus, der inducerer migrationen af CL fra huden til lymfeknuderne og produktionen af CL TNF A er interaktionen mellem CD40 CL -molekylet med CD40L hud T -celler. TNF a såvel som IL-1 b er de vigtigste faktorer, der er ansvarlige for implementeringen af denne proces; IL-4 (gennem undertrykkelse af Rc-ekspression for TNF a) og IL-10 undertrykker det. Den indledende fase af migration - frigivelsen af CL til dermis - tilvejebringes af virkningen af GM-CSF.
Det er almindeligt accepteret, at i processen med migration til lymfeknuderne gennem de afferente lymfekar, gennemgår CL'er differentiering, hvilket resulterer i en svækkelse af deres evne til at binde og behandle Ag og en stigning i antigenpræsenterende evne (fig. 39). . Et mellemtrin i transformationen af CL til modne dendritiske (interdigiterende) celler er slørede lymfeceller. TNF b undertrykker CL-differentiering. Lokaliseringen af modne CL'er (interdigiterende celler) i lymfeknudernes thymus-afhængige zoner afhænger af cellernes sekretion fra mikromiljøet af disse zoner (inklusive de interdigiterende celler selv) af b-kemokiner CCL19 (ELC) og især CCL21 (SLC), der interagerer med Rc CCR7 CL. CL's vej fra huden til den regionale lymfeknude og lokalisering i den blev sporet i detaljer i forbindelse med opdagelsen af en specifik markør for disse celler (fraværende på andre dendritiske celler), som viste sig at være langerin (CD208). Under betingelser med en inflammatorisk reaktion reduceres opholdstiden for LA i epidermis fra 18 måneder til 2 uger; samtidig stimuleres tilstrømningen af CCR2+ – forstadier til LA fra kredsløbet som reaktion på udskillelsen af b-kemokin MCP-1 fra keratinocytter.
Ris. 39 . Karakterisering af membranfænotypen og antigenpræsenterende aktivitet af huddendritiske celler på forskellige stadier af deres modning og migration til lymfeknuderne. Over stregerne i toppen er cytokiner, der fremmer frigivelsen af Langerhans-celler til lymfen (venstre) og kemokiner, der sikrer lokaliseringen af interdigiterende celler i lymfeknudens thymus-afhængige zoner [højre); GM-CSF - granulocyt-makrofag kolonistimulerende faktor; CCR - Rc p-kemokiner (fra CC-kemokinreceptor); CLA - Ag af hudlymfocytter (fra kutan lymfocytantigen); MCP - Hæmatractant protein af monocytter (fra - Monocyte Chemoattractant Protein); SLC - Hæmokin af sekundært lymfoidt stof (fra - Secondarylymphoid vævskemokin); VLA er en meget sen aktiverings-AG (fra - Very Late Activation Antigen).
Det antages således, at CLS udfører funktionen af APC'er, men processen med at implementere denne funktion er fordelt i tid og rum: i huden, CLS endocytize og proces AG, og i den regionale lymfeknude, hvor de vandrer, præsenterer CLS en antigent peptid til T-hjælpere, herunder således et specifikt immunrespons. Aktiviteten af CL manifesterer sig i form af induktion af en T-celle-respons i enhver form for immunrespons, men det er især tydeligt i tilfælde af beskyttelse mod vira, der ændrer overfladen af keratinocytter, melanocytter og andre celler omkring CL . Det ligger også til grund for udviklingen af kontaktoverfølsomhed, da allergener, herunder lav molekylvægt, kan vise deres sensibiliserende virkning først efter binding til MHC -klasse II -molekyle på overfladen af CL og den efterfølgende udvikling af en allergisk reaktion i henhold til det sædvanlige mekanisme. In vitro denne reaktion kan kun reproduceres ved hjælp af CL, men ikke andre celler. Tilstedeværelsen af CL i hudtransplantater er en forudsætning for deres afvisning i tilfælde af forskelle i MHC klasse II gener. I dette tilfælde tjener CL'er som "passagerleukocytter", som er obligatoriske for at udløse en afvisningsreaktion.
Det er blevet fastslået, at CL'er er meget følsomme over for ultraviolet bestråling af huden. Efter bestråling forårsager virkningen af sensibiliserende stoffer (for eksempel dinitrophenol) på epidermis ikke kun ikke kontakt overfølsomhed, men inducerer også specifik manglende reaktion på dette allergen. Dette skyldes det faktum, at CLS under påvirkning af ultraviolet lys inaktiveres, og i stedet for dem opfatter antigener APC'er, der aktiverer overvejende T -undertrykkere - direkte (Granstein -celler i mus) eller indirekte gennem aktivering af inducere af CD4, CD45RA fænotype suppressor (makrofager af CD1 fænotypen - , HLA - DR + person).
CL er således celler af knoglemarvsoprindelse, der tilbringer en del af deres livscyklus i huden. Når de udsættes for skadelige faktorer, infektionsmidler, allergener og andre faktorer, aktiveres CLS og forlader huden, der migrerer til den regionale lymfeknude. I migrationsprocessen modnes de til stadiet af aktive APC'er og en gang i lymfeknuden præsenterer et antigen peptid til T-Helpers og udløser derved en specifik immunrespons.
Det er gratis at sende annoncer, og registrering er ikke nødvendig. Men der er præ-moderering af annoncer.
Langerhans celler
Langerhans-celler, opkaldt efter deres opdager - P. Langerhans, er i øjeblikket anerkendt som en undertype af dendritiske celler (indtil for nylig blev de klassificeret som vævsmakrofager i huden). Disse celler findes i epitelvæv, de er i stand til fagocytose, udtrykker ikke B7-costimulatoren og spiller en væsentlig rolle i at give kroppens immunrespons.
Mekanismen for deres funktion er som følger. Langerhanske celler migrerer gennem lymfekarrene til de nærmeste lymfeknuder, hvor de omdannes til standard dendritiske celler med overflade B7 co-receptorer. Dette bidrager til skabelsen af betingelser for inklusion af immunresponser af CD8 T- og CD4 T-celler i tilfælde af penetration af patogener gennem beskadigede områder af huden og sikrer isolering af mikroorganismer. Langerhans-celler er også involveret i at give et immunrespons i nærvær af kontaktoverfølsomhed i huden. Derudover er de i stand til at bevare antigener i lang tid, flytte dem til lymfeknuderne og opretholde immunologisk hukommelse.
Langerhans-celler har en endokrin funktion, der sikrer udskillelsen af mange stoffer, der er nødvendige for epidermis' levetid (især prostaglandiner, gamma-interferon, interleukin-1 samt elementer, der regulerer celledeling og proteinsyntese). Der er videnskabelige data om den særlige antivirale virkning af Langerhans-celler og om deres deltagelse i ødelæggelsen af papillomer.
Det er blevet fastslået, at i processen med kroniske sygdomme, ældning af kroppen, UV-bestråling samt forgiftning reduceres antallet af Langerhans-celler betydeligt.
Markedsanalyse
- Globalt kosmetikemballagemarked – digitalt fokus og bæredygtighed
- Global Cosmetics Market Review 2018 Det bedste år i historien på det globale skønhedsmarked gennem de sidste 20 år
- Kosmetiske nyheder fra 2018 eller skønhedsindustrien i et nyt format
Skønhedssaloner i Jekaterinburg Skønhedssaloner i Novosibirsk
Seneste blogindlæg på vores side
- Naturcreme / Havtornolie - "flydende guld" til ansigtet
- Naturcreme / Skin Superfood fra Navarra
- Naturcreme /
Celler - få en fungerende rabatkupon hos Akademika til Kosmetikgalleriet eller profitable celler at købe med gratis fragt på udsalg i Kosmetikgalleriet
Langerhans dendritiske celler- Orale og genitale slimhindeceller Bioteknologiske emner EN dendritiske Langerhans-celle …
langerhans øer- grupper af celler i bugspytkirtlen hos mennesker og hvirveldyr (undtagen cyclostomer), der danner dens intrasekretoriske del; udskiller hormonerne insulin og glukagon i blodet. Opkaldt efter den tyske videnskabsmand P. Langerhans (P. ... ... Store sovjetiske encyklopædi
Langerhans øer- Må ikke forveksles med Langerhans-celler indeholdt i epidermale væv i bioyal ... Wikipedia
delta celler- δ-celler (eller D-celler) celler, der producerer hormonet somatostatin; de findes i maven, tarmene og også i de Langerhanske øer i bugspytkirtlen. Hos gnavere er deltaceller hovedsageligt placeret langs øernes periferi, i ... ... Wikipedia
holmen Langerhans- Langerhans (pancreas) ø Langerhans. Grupper af celler i bugspytkirtelvæv (hos alle hvirveldyr undtagen cyclostomer) alfaceller O.L. udskiller glukagon
holmen Langerhans- Grupper af celler i bugspytkirtelvæv (hos alle hvirveldyr undtagen cyclostomer) alfaceller O.L. udskiller glukagon og betaceller udskiller insulin; O.L. størrelser 50 500 mikron. [Arefiev V.A., Lisovenko L.A. Engelsk russisk forklarende ordbog over genetisk ... ... Teknisk oversætterhåndbog
Antigenpræsenterende celler- Immunokompetente celler, der er ansvarlige for behandlingen (behandling) af antigen og dets efterfølgende præsentation for forskellige populationer af lymfocytter. A.k. omfatter især makrofager, interdigitante celler i den paracorticale zone af lymfeknuderne, som dannes ... ... Mikrobiologi ordbog
Langerhans øer- små klynger af celler spredt ud over bugspytkirtlen, der udskiller hormonerne insulin og glukagon. Der er tre histologiske typer af disse celler: alfa-, beta- og deltaceller; de producerer henholdsvis glukagon, insulin og ... ... medicinske termer
ØER AF LANGERHANS- (Langerhanske øer) små klynger af celler spredt ud over bugspytkirtlen, der udskiller hormonerne insulin og glukagon. Der er tre histologiske typer af disse celler: alfa-, beta- og deltaceller; de producerer henholdsvis ...... Forklarende ordbog for medicin
Immunitet- I Immunitet (lat. immunitas frigivelse, at slippe af med noget) kroppens immunitet over for forskellige infektionsstoffer (vira, bakterier, svampe, protozoer, helminths) og deres stofskifteprodukter, samt over for væv og stoffer ... ... Medicinsk encyklopædi
I det 19. århundrede opdagede en ung videnskabsmand fra Tyskland uensartetheden af bugspytkirtelvæv. Celler, der adskilte sig fra hovedmassen, var placeret i små klynger, øer. Grupper af celler blev senere opkaldt efter patologen - Langerhanske øer (OL).
Deres andel i det samlede volumen af væv er ikke mere end 1-2%, men denne lille del af kirtlen udfører sin funktion, forskellig fra fordøjelseskanalen.
Предназначение островков Лангерганса
 Hoveddelen af cellerne i bugspytkirtlen (PZh) producerer enzymer, der fremmer fordøjelsen. Funktionen af ø-klynger er anderledes - de syntetiserer hormoner, så de omtales som det endokrine system.
Hoveddelen af cellerne i bugspytkirtlen (PZh) producerer enzymer, der fremmer fordøjelsen. Funktionen af ø-klynger er anderledes - de syntetiserer hormoner, så de omtales som det endokrine system.
Således er bugspytkirtlen en del af to hovedkroppssystemer - fordøjelsessystemet og det endokrine. Островки являются микроорганами, вырабатывающими 5 видов гормонов.
De fleste af bugspytkirtelgrupperne er placeret i bugspytkirtlens hale, selvom kaotiske mosaikindeslutninger fanger hele det eksokrine væv.
OL er ansvarlige for reguleringen af kulhydratmetabolismen og understøtter arbejdet i andre endokrine organer.
Histologisk struktur
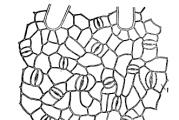
Hver ø er et selvfungerende element. Tilsammen udgør de en kompleks øgruppe, der består af individuelle celler og større formationer. Deres størrelse varierer betydeligt, fra en endokrin celle til en moden, stor ø (>100 µm).
I pancreasgrupper er der bygget et hierarki af cellearrangementer, der er 5 typer af dem, alle udfører deres rolle. Hver ø er omgivet af bindevæv, har lobuler, hvor kapillærer er placeret.
Grupper af betaceller er placeret i midten, langs kanterne af formationerne er alfa- og deltaceller. Чем больше размер островка, тем больше в нем периферийных клеток.
Øerne har ikke kanaler, de producerede hormoner udskilles gennem kapillærsystemet.

celletyper
Forskellige grupper af celler producerer deres egen type hormon, der regulerer fordøjelsen, lipid- og kulhydratmetabolismen.
- Alfa celler. Denne gruppe af OL'er er placeret langs kanten af øerne; deres volumen er 15-20% af den samlede størrelse. De syntetiserer glukagon, et hormon, der regulerer mængden af glukose i blodet.
- beta-celler. Группируются в центре островков и составляют большую часть их объема, 60- 80%. De syntetiserer insulin, omkring 2 mg om dagen.
- delta celler. Er ansvarlige for produktionen af somatostatin, fra 3 til 10%.
- Epsilon celler. Mængden fra den samlede masse er ikke mere end 1%. Deres produkt er ghrelin.
- PP celler. Hormonet pancreaspolypeptid produceres af denne del af Ol. De udgør op til 5% af holmene.
I løbet af livet falder andelen af den endokrine komponent i bugspytkirtlen - fra 6% i de første levemåneder til 1-2% ved 50 års alderen.
Hormonel aktivitet
Bugspytkirtlens hormonelle rolle er stor.
De aktive stoffer syntetiseret i små øer leveres af blodbanen til organerne og regulerer metabolismen af kulhydrater:
- Insulinets hovedopgave er at minimere blodsukkeret. Det øger absorptionen af glucose af cellevægge, fremskynder dets oxidation og hjælper med at lagre det som glykogen. Overtrædelse af syntesen af hormonet fører til udvikling af type 1-diabetes. Samtidig viser blodprøver tilstedeværelsen af antistoffer mod dyrlægeceller. Type 2-diabetes udvikler sig, når væv bliver mindre følsomme over for insulin.
- Glukagon udfører den modsatte funktion - det øger sukkerniveauet, regulerer produktionen af glukose i leveren og fremskynder nedbrydningen af lipider. To hormoner, der supplerer hinandens virkning, harmoniserer indholdet af glukose - et stof, der sikrer kroppens vitale aktivitet på celleniveau.
- Somatostatin bremser mange hormoner. I dette tilfælde er der et fald i absorptionshastigheden af sukker fra mad, et fald i syntesen af fordøjelsesenzymer og et fald i mængden af glukagon.
- Pancreatisk polypeptid reducerer mængden af enzymer, bremser frigivelsen af galde og bilirubin. Det menes, at det stopper forbruget af fordøjelsesenzymer og holder dem indtil næste måltid.
- Grelin betragtes som hormonsult eller mæthed. Dens produktion giver et signal til kroppen om en følelse af sult.
Mængden af producerede hormoner afhænger af glukose modtaget fra fødevarer og hastigheden af dets oxidation. Med en stigning i mængden øges insulinproduktionen. Syntese udløses ved en koncentration på 5,5 mmol/l i plasma.
Fremprovokere produktionen af insulin kan ikke kun spise. Hos en sund person noteres den maksimale koncentration i en periode med stærk fysisk stress, stress.
Den endokrine del af bugspytkirtlen producerer hormoner, der har en afgørende effekt på hele kroppen. Patologiske ændringer i OB kan forstyrre funktionen af alle organer.
Video om insulinets opgaver i den menneskelige krop:
Nederlaget for den endokrine del af bugspytkirtlen og dens behandling
Årsagen til OL-skader kan være genetisk disposition, infektioner og forgiftninger, inflammatoriske sygdomme, immunproblemer.
Som et resultat er der et ophør eller et betydeligt fald i produktionen af hormoner af forskellige celler i øerne.
Som et resultat kan det udvikle sig:
- SD type 1. Det er karakteriseret ved fravær eller mangel på insulin.
- SD type 2. Det bestemmes af kroppens manglende evne til at bruge det producerede hormon.
- Svangerskabsdiabetes udvikler sig under graviditeten.
- Andre typer af diabetes mellitus (MODY).
- neuroendokrine tumorer.
Hovedprincipperne for behandling af type 1-diabetes mellitus er indførelse af insulin i kroppen, hvis produktion er svækket eller reduceret. Der er to typer insulin - hurtigtvirkende og langtidsvirkende. Sidstnævnte type efterligner produktionen af bugspytkirtelhormon.
 Type 2-diabetes kræver en streng diæt, moderat motion og sukkerforbrændende medicin.
Type 2-diabetes kræver en streng diæt, moderat motion og sukkerforbrændende medicin.
Diabetes er stigende på verdensplan og bliver allerede kaldt det 21. århundredes pest. Derfor leder medicinske forskningscentre efter måder at bekæmpe sygdomme på øerne i Langerhans.
Processer i bugspytkirtlen udvikler sig hurtigt og fører til øernes død, som burde syntetisere hormoner.
I de senere år er det blevet kendt:
- stamceller transplanteret til bugspytkirtelvævet slår godt rod og er i stand til at producere hormonet i fremtiden, da de begynder at fungere som betaceller;
- OL producerer flere hormoner, hvis en del af bugspytkirtlens kirtelvæv fjernes.
Dette giver patienter mulighed for at opgive den konstante brug af stoffer, en streng diæt og vende tilbage til en normal livsstil. Problemet er fortsat immunsystemet, som kan afvise transplanterede celler.
En anden mulig behandling er transplantation af en del af øvævet fra en donor. Denne metode erstatter installationen af en kunstig bugspytkirtel eller dens fuldstændige transplantation fra en donor. Samtidig er det muligt at stoppe udviklingen af sygdommen og normalisere blodsukkeret.
Der er gennemført vellykkede operationer, hvorefter patienter med type 1-diabetes ikke længere behøver at administrere insulin. Kroppen genoprettede populationen af betaceller, syntesen af dets eget insulin blev genoptaget. Postoperativt blev immunsuppressiv terapi administreret for at forhindre afstødning.
Videomateriale om funktionerne af glukose og diabetes:
Medicinske institutter arbejder på at undersøge muligheden for at transplantere en bugspytkirtel fra en gris. De første lægemidler til behandling af diabetes brugte bare dele af bugspytkirtlen hos grise.
Forskere er enige om, at det er nødvendigt at studere de strukturelle træk og arbejde fra Langerhans øer på grund af det store antal vigtige funktioner, som hormonerne syntetiserede i dem udfører.
Det konstante indtag af kunstige hormoner hjælper ikke med at besejre sygdommen og forværrer patientens livskvalitet. Nederlaget for denne lille del af bugspytkirtlen forårsager dybtgående forstyrrelse af hele kroppen, så forskningen fortsætter.